
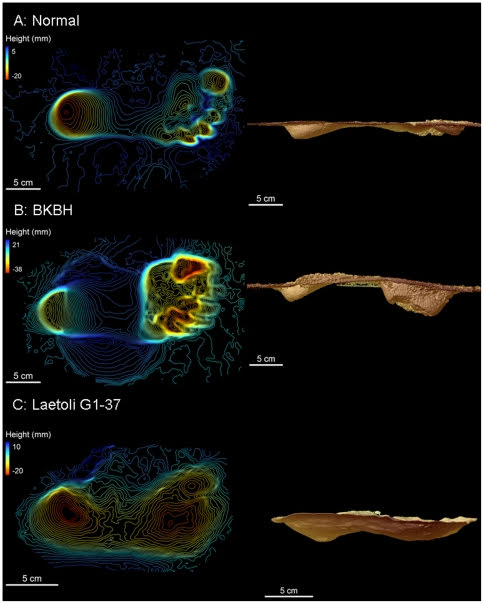
今回は、上肢帯の起源である「肩帯」について解説していく。
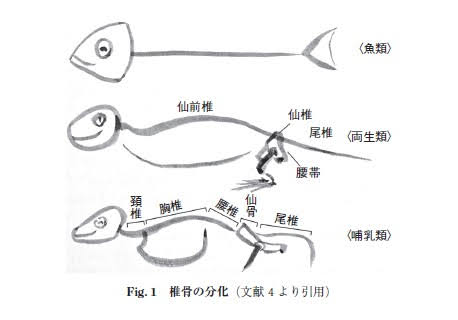
脊椎動物の起源は、ナメクジウオが属する頭索動物であるという説が有力であり、その頭索動物から魚類が誕生した。
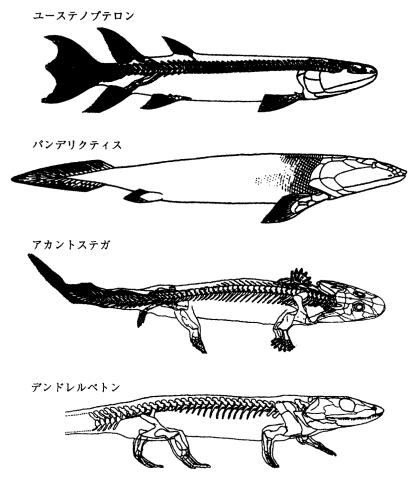
魚類の鰭が陸上生活で進化してしたものが手や腕となった。
上肢帯の基部を構成する部分を「肩帯」と呼び、以下の骨で構成される。
肩甲骨
上肩甲骨
前烏口骨
烏口骨
上鎖骨(擬鎖骨)
上上鎖骨(上擬鎖骨)
鎖骨
間鎖骨
ここで挙げた骨は特定の動物群にしか現れない物や、進化の過程で消滅したりする物が含まれており、これら全てを同時に持つ脊椎動物はいない。
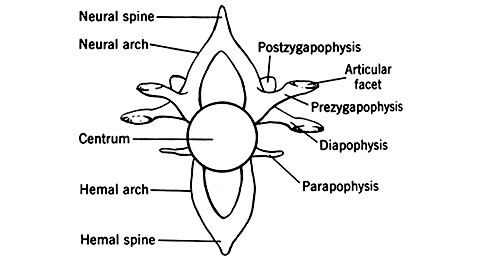
間鎖骨以外は左右対である。(1)
まずは魚類の肩帯について紹介する。
コイ科魚類の骨格(2)
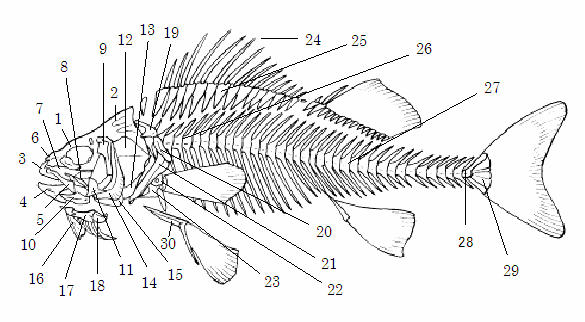
上擬鎖骨(20)擬鎖骨(21)肩甲骨(22)烏口骨(23)
頭部側面観(3)
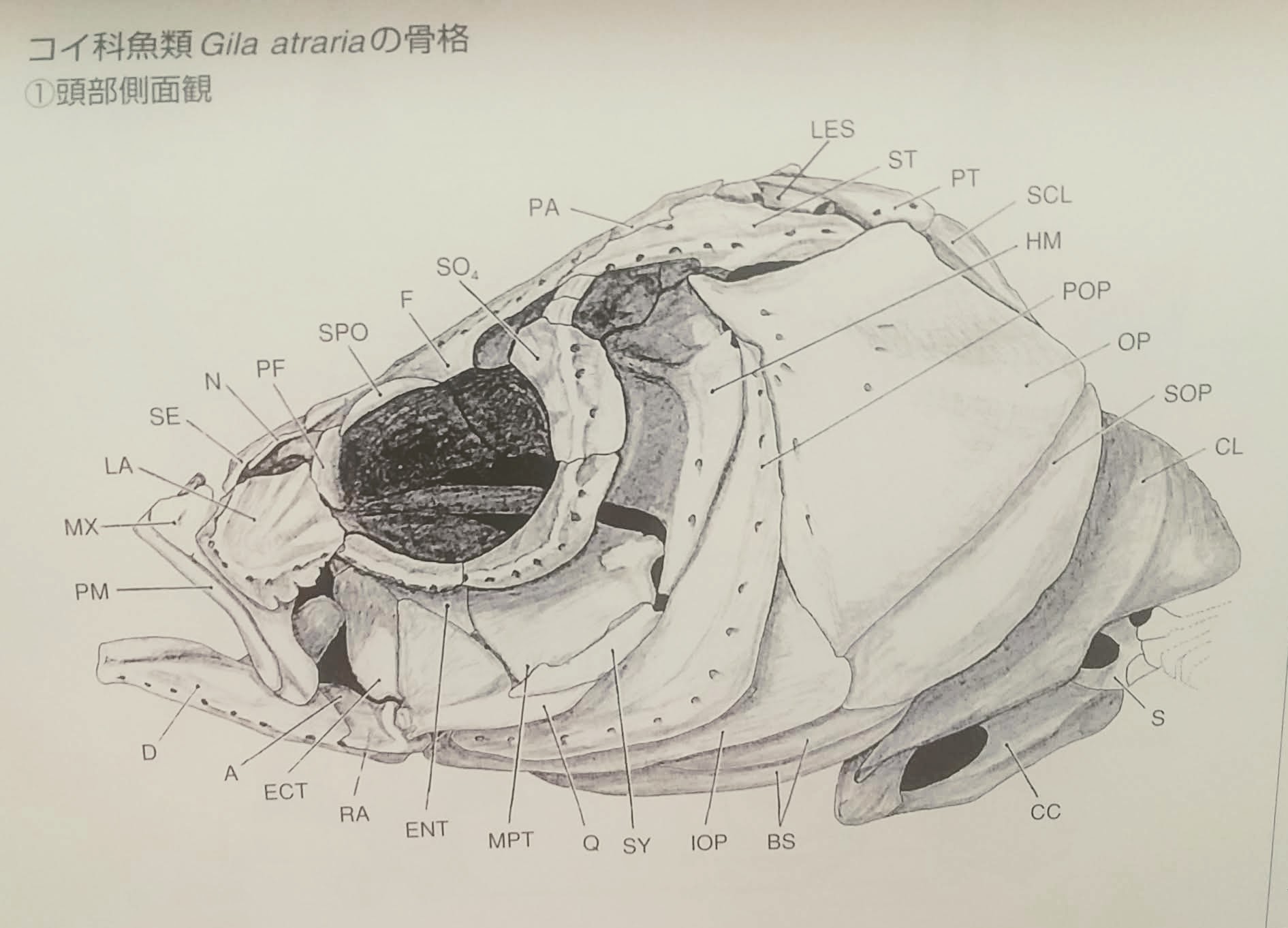
烏口骨(CC)擬鎖骨(CL)肩甲骨(S)上擬鎖骨(SCL)
肩帯とそのいろいろな骨(3)
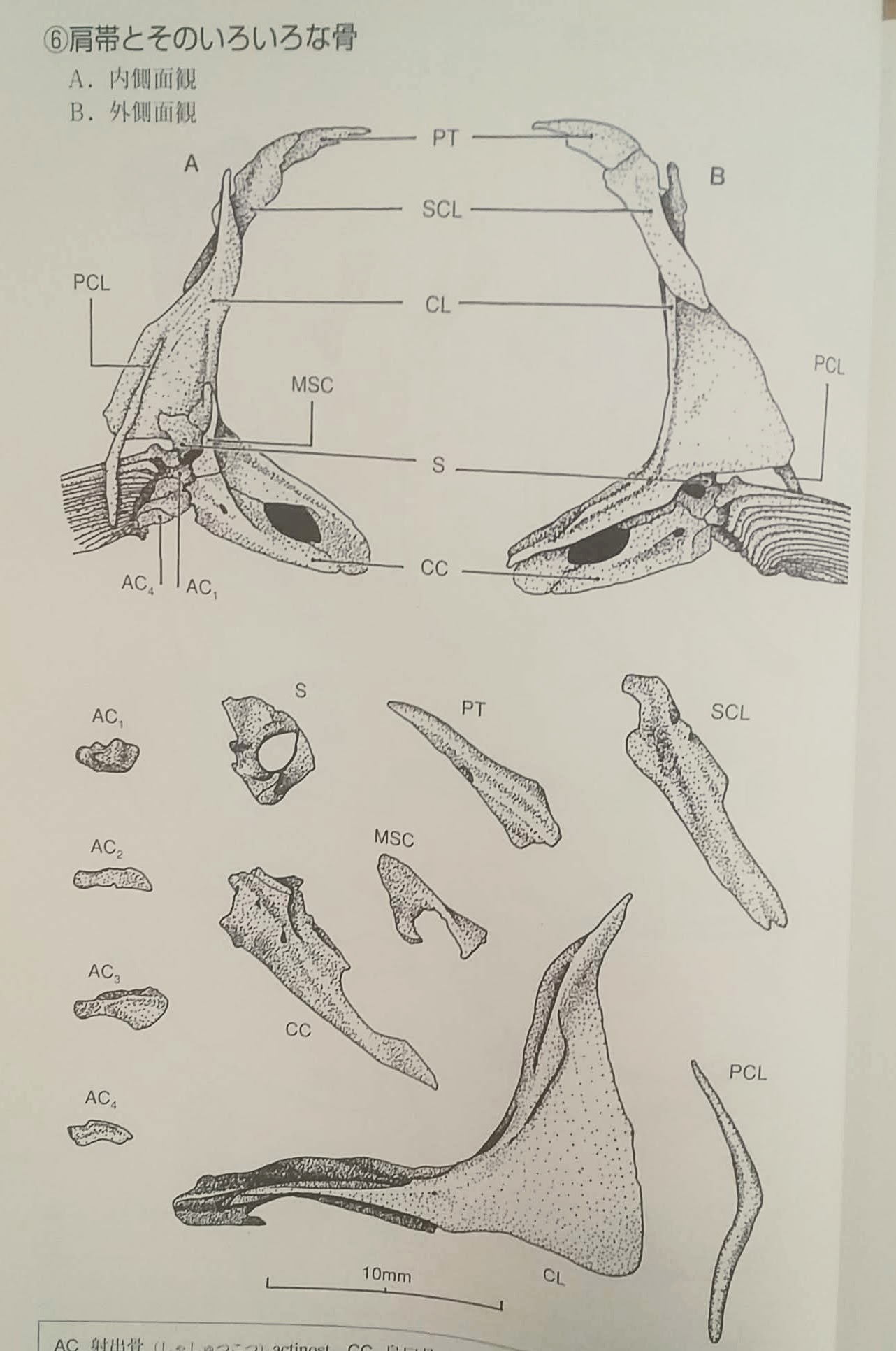
後側頭骨(PT)上擬鎖骨(SCL)擬鎖骨(CL)肩甲骨(S)後擬鎖骨(PCL)烏口骨(CC)
肩帯を構成する骨において、鎖骨群は頭蓋骨後縁部から派生した骨であるため、頭蓋骨と同じく皮骨性骨格である。
その他の骨は内骨格性骨格である。
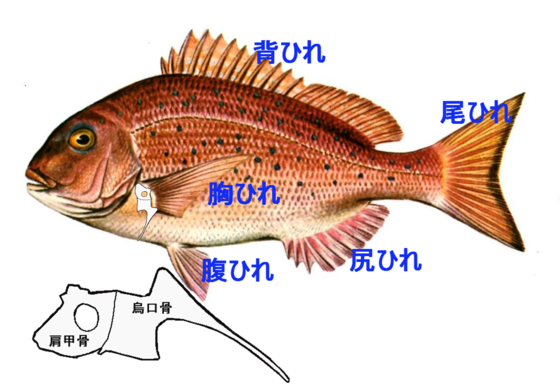
一部の魚には、俗に「鯛の鯛」と呼ばれる部位がある。
これは胸鰭を構成する肩帯の形がその魚に似ていることが名前の由来とされている。(4)(5)
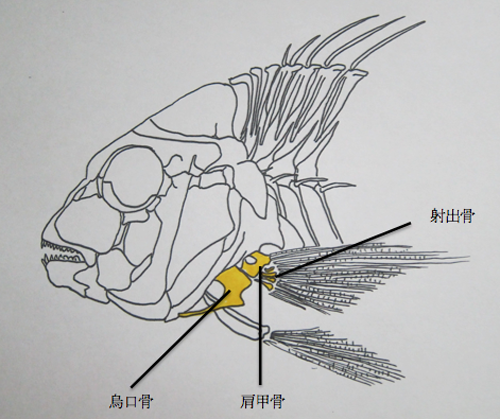

肩甲骨は、鰓裂後方部を構成する骨から派生したものと考えられている。
初期の脊椎動物である魚類の肩帯は、ただ胸鰭を動かすためのものであった。
魚類から両生類への進化で肩帯に起こった最も重要なことは、上上鎖骨が消失したことにより、肩帯が完全に頭蓋から切り離されたことである。
これにより、前肢は自由度が増し、さらに”頚部”が生まれる要因にもなった。(1)
鎖骨群は頭部から切り離された後下方へ下がり、間鎖骨によって左右の鎖骨は連結されることになる。
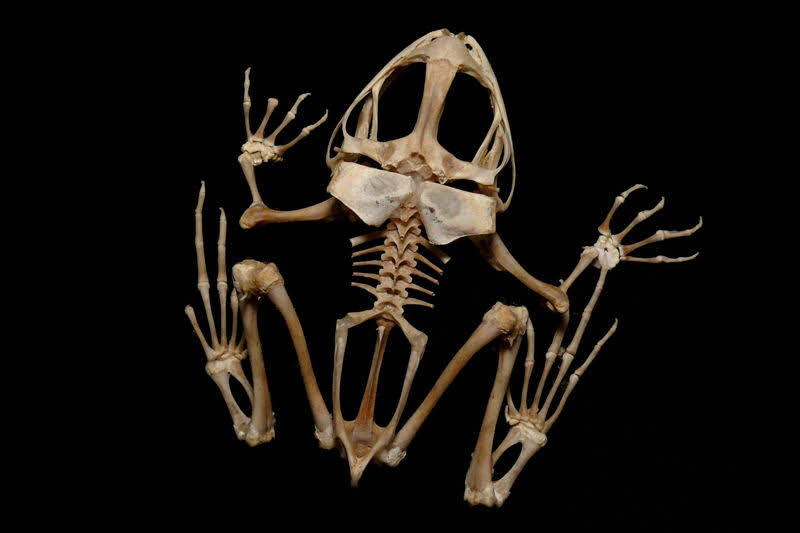
ヒキガエルの肩帯(1)
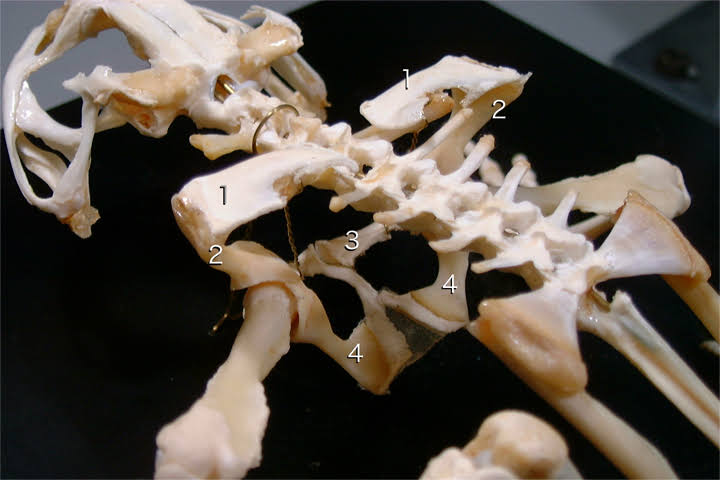
1.上肩甲骨 2.肩甲骨 3.鎖骨 4.前烏口骨
魚類では、肩帯は胸鰭を動かす為だけに存在しているが、両生類になると体を支持するという機能も合わせ持つようになった。
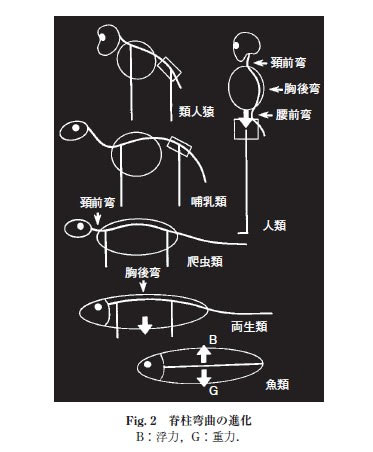
陸上では水中程方向転換が容易ではないことから、頸部の自由度が必要になったと想像出来る。
肩帯は頭部からを分離することで、その自由度とロコモーション機能を高めることが出来た。
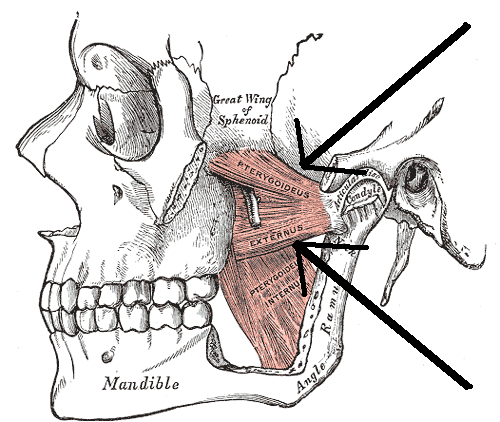
爬虫類になると、前烏口骨の後方に新たな化骨中心が現れ、それが烏口骨となった。
さらにほとんどの進化系列で上鎖骨は退化傾向を示し、最終的に消失する。
主竜類の系統が内骨格性肩帯として肩甲骨と前烏口骨が基本となるのに対し、単弓類の系統では前烏口骨が退化していき、肩甲骨と烏口骨が内骨格性肩帯の主要な構成要素となっていく。
現生の爬虫類でいうと、肩甲骨・前烏口骨・鎖骨・間鎖骨が肩帯の基本的な構成骨である。
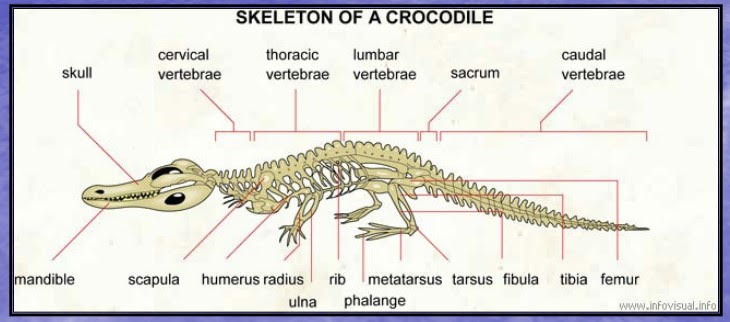
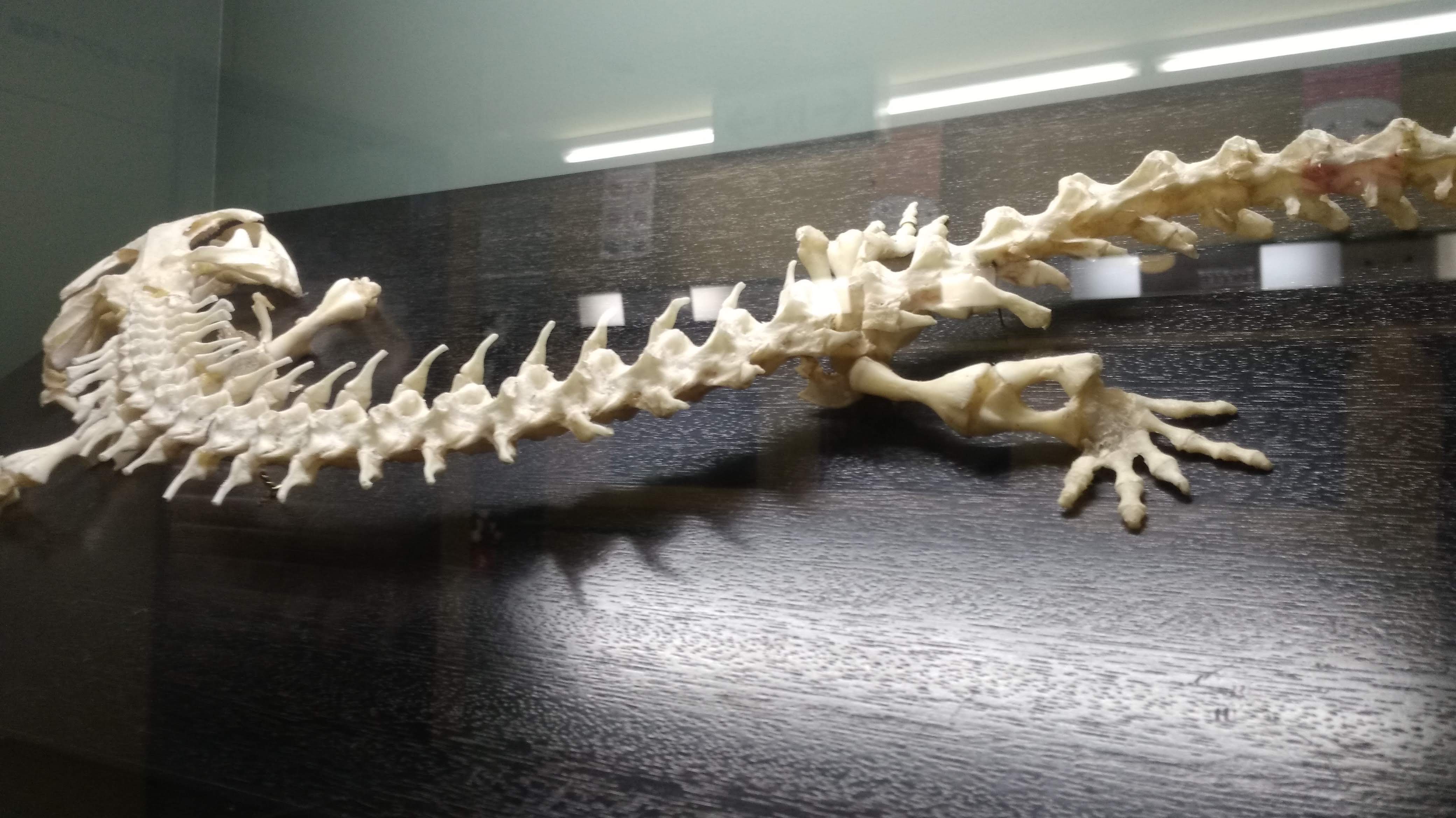
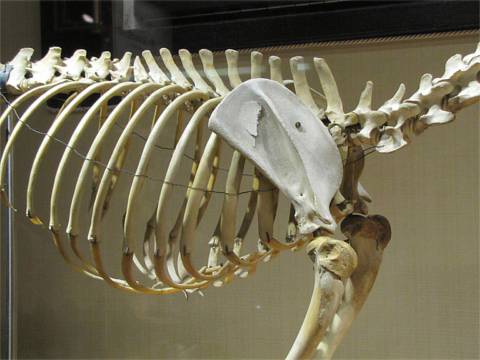
ただし、主竜類は鎖骨も退化の傾向を示すので、ワニ類には鎖骨はない。(1)
ワニの骨格(6)

爬虫類は哺乳類の祖先ではなく、両生類から派生した単弓類が哺乳類の祖先である。
両生類から別に派生した爬虫類から恐竜、鳥類が派生し、哺乳類は別系統の単弓類から派生したのである。
鳥類の基本的な構成は、爬虫類の主竜類と同じである。
ただし左右の鎖骨(とおそらく間鎖骨も)は癒合してV字またはY字形となっており、叉骨 (furcula) または暢思骨 (wishbone) という名で呼ばれることがある。
肩甲骨は薄く細長くなって伸張し、脊椎とおおむね平行になっている。
前烏口骨も棒状に伸張するが、肩甲骨とは異なり強く頑丈になっている。
これは巨大な胸骨に起始し上腕骨に至る強大な飛翔筋が収縮する際の荷重に耐えるためであると考えられている。(1)
ハトの骨格(7)
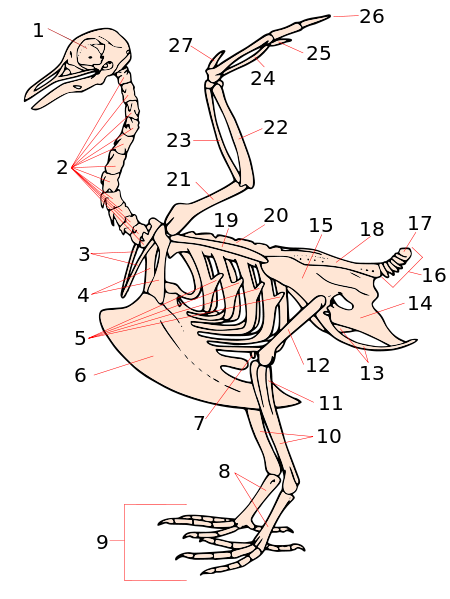
叉骨(3)烏口骨(4)竜骨突起(6)肩甲骨(19)
単弓類の盤竜類段階で持っていた肩帯構成骨は、肩甲骨・前烏口骨・烏口骨・上鎖骨・鎖骨・間鎖骨の6種であった。
しかし、そこから獣弓類を経て哺乳類に進化した際に失った骨は上鎖骨のみである。
単孔類のカモノハシは肩甲骨・前烏口骨・烏口骨・鎖骨・間鎖骨の5種の骨からなる肩帯を保持している。
現在のヒトを含む真獣類が持っている肩帯構成骨は肩甲骨・鎖骨の2種にまで減少しており、単孔類の肩帯が原始的であるという言われ方をするのはそのためである。
その後の進化の中で、間鎖骨と前烏口骨は消失し、烏口骨は肩甲骨と癒合して烏口突起 (coracoid process) と呼ばれる部位になった。
烏口突起とはヒトではその名の通り鳥のくちばしのように鍵形に曲がった突起であり、むしろ烏口骨という名称が烏口突起に由来している。
それとほぼ時を同じくして、それまでの肩甲骨の前縁から更に前方に筋の付着面が形成され、その新しい付着面を棘上窩、それまで前縁だった部分を肩甲棘、それまでの筋付着面を棘下窩と呼ぶようになった。
最終的には、真獣類の肩帯の内骨格性成分は肩甲骨のみとなっている。一方、皮骨性骨格成分として最後に残ったのは鎖骨である。鎖骨は上腕を様々な方向に回転させる樹上性の動物ではよく発達しているが、有蹄類・食肉類などを始めとした多くの群で消失している。
これは、走行・跳躍など前肢の前後への運動が主となる場合には、肩甲骨遠位部の自由度を大きくしておいた方がよいためであろうと解釈されている。
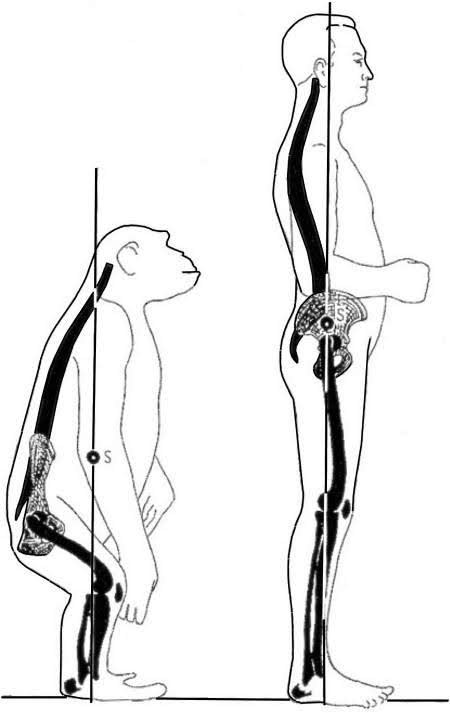
ヒトの肩帯(1)
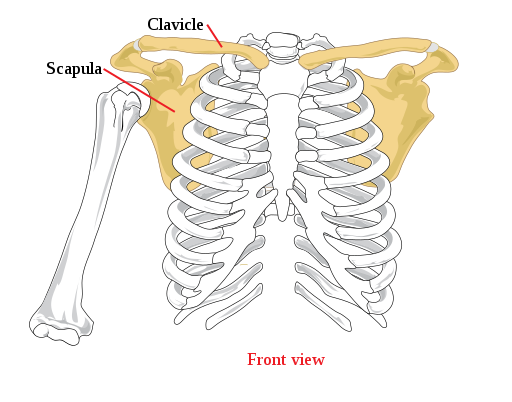
哺乳類でもウマ、イヌ、ウシ、ゾウの様な走行性の哺乳類等では退化している場合も多い。
木登りをするネコには鎖骨が存在するが、木登りをしないチーターには鎖骨は無い。
鎖骨がないといわゆる抱きつく所作(前脚を内側に曲げ保持すること)が困難で鎖骨のない動物は木登りができないことから、早期に草原に進出した動物は長距離移動に適応して鎖骨が退化し、長期間森林に生息した動物には鎖骨が残っているのではないかと考えられている。(1)
ネコの鎖骨(8)
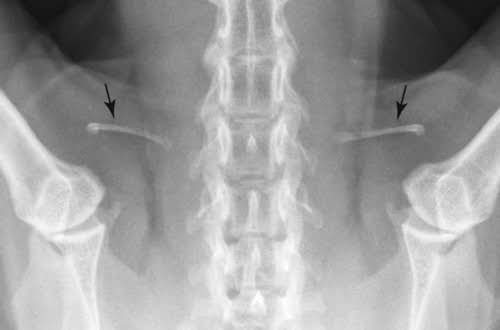
鎖骨が残存する必要性は挙上位での上肢のコントロールであり、樹上で生活をする霊長類の鎖骨は退化していない。
ナチュラリゼーションにおける『這うワーク』は、魚の胸鰭から進化した両生類の上肢の動きをモチーフにしており、その上肢の原始的な機能を取り戻す意味があると言えるでしょう。
参考文献
(1)ウィキペディア,https://ja.wikipedia.org/wiki/肩帯,2019年9月27日アクセス
(2)上野輝彌・坂本一男(1999)『魚の分類の図鑑』 東海大学出版会
(3)上野輝彌・坂本一男(2005)『新版 魚の分類の図鑑』 東海大学出版会
(4)落合明編(1994)『魚類解剖大図鑑』 緑書房
(5)高知新聞朝刊(2013年2月2日) 表情?豊かな鯛の鯛
(6)Wikipedia Crocodilian armor,https://en.wikipedia.org/wiki/Crocodilian_armor2019年6月19日アクセス.
(7)ウィキペディア,https://ja.wikipedia.org/wiki/鳥類の体の構造,2019年9月27日アクセス
(8)Radiographic Signs of Joint Disease in Dogs and Cats,https://veteriankey.com/radiographic-signs-of-joint-disease-in-dogs-and-cats/,2019年9月27日アクセス






{kind=link}
{kind=link}
{kind=link}